
英语原文共 10 页,剩余内容已隐藏,支付完成后下载完整资料
对ISCR的研究揭示了金属依赖性调控DNA结合特异性的独特机制
SenapathyRajagopalan,莎拉.杰特,PetrusHZwart,RichardGBrennan,凯文.J.菲利普斯,帕特里夏.杰.凯利
来自大肠杆菌的ISCR是一种不寻常的金属调节剂,因为APO和铁硫(Fe-S)-ISCR都调节转录,并表现出不同的DNA结合特异性。在这里,我们报告了ISCR的结构和生化研究,表明Fe-S连接后蛋白质-DNA界面的重塑拓宽了ISCR的DNA结合特异性,从仅将2型模体结合到1型和2型模体。对具有松弛靶位判别的apo-IscR变体的分析确定了野生型apo-IscR中的一个关键残基,我们建议,与1型模体进行不利的相互作用。在Fe-S结合时,这些相互作用显然被移除,从而允许ISCR结合1型和2型模体。这些数据表明配体介导的DNA位点识别的独特机制,即金属簇连接重新定位蛋白质特异性决定簇以扩大DNA靶位点选择,从而允许holo-IscR的更广泛的转录反应。
Fe-S蛋白是一种古老的蛋白质,存在于生命的各个王国。它们在基因组维持、转录、翻译和代谢中具有关键的结构、催化或电子转移作用。一些最近发现的调节蛋白利用Fe-S簇作为各种小分子的传感器,利用这些金属中心的多功能化学反应活性。研究良好的调节剂[4Fe-4S]-FNR的调节反应是大多数金属调节因子的典型,其中同源金属中心的结合将转录因子从非活性状态切换到活性状态,并相应地改变其调节因子的表达。相比之下,大肠杆菌[2Fe-2S]转录因子ISCR已成为一种不寻常的Tran转录调节剂,由于其无簇形式(APO-ISCR)和[2Fe-2S]结合形式都是活性转录调节剂。簇连接明显改变了DNA结合的特异性,从而改变了IscR控制的基因集。
ISCR在细菌中广泛保守是大的Rrf2家族的一部分,有翼螺旋-转螺旋(WHTH)转录因子。它首先被发现为ISC操纵子的负自调节因子,编码ISCR和ISCFe-S生物发生途径的成员。然而,现在已经知道IscR是一个直接或间接控制40个基因表达的全球调节因子。对ISCR调控基因的分析表明,ISCR识别1型和2型DNA结合模体,这取决于[2Fe-2S]簇是否与蛋白质连接。只有[2Fe-2S]-IscR结合1型位点,而[2Fe-2S]-ISCR和APO-ISCR都结合了具有相似高亲和力的2型位点表示Fe-S簇不需要与2型位点交互。的IscR辨别能力在两个DNA模体之间是出乎意料的,因为ISCR的预测结构只包含一个wHTHDNA结合结构域,而其他已知的含有Fe-S簇的转录因子(例如FNR和SoxR)在缺乏金属簇时似乎不能调节转录。在[2Fe-2S]簇连接时,IscR靶位识别的特征和改变IscRDNA结合特异性的结构变化是未知的。
为了深入了解IscR如何识别两种不同的DNA模体,我们使用了X射线晶体学和结构导向突变。我们结晶了一个缺乏Fe-S簇的IscR突变体,它与编码氢化酶-1的hya操纵子启动子的一个研究良好的2型位点结合。根据IscR-hya的结构
复合物,我们在介导蛋白质-DNA相互作用的关键残基处进行丙氨酸替换,并测定每个蛋白质变体的[2Fe-2S]结合和载脂蛋白形式结合1型和2型位点的能力。我们发现Glu43的侧链是一个主要的鉴别器,允许ApoISCR区分1型和2型结合位点。这些研究提示了一种以前未知的DNA结合转录因子的金属依赖性调控机制,其中DNA结合特异性明显被金属辅因子连接后的单键侧链的重新连接所拓宽。
结果
与2型DNA位点结合的apo-IscR的结构
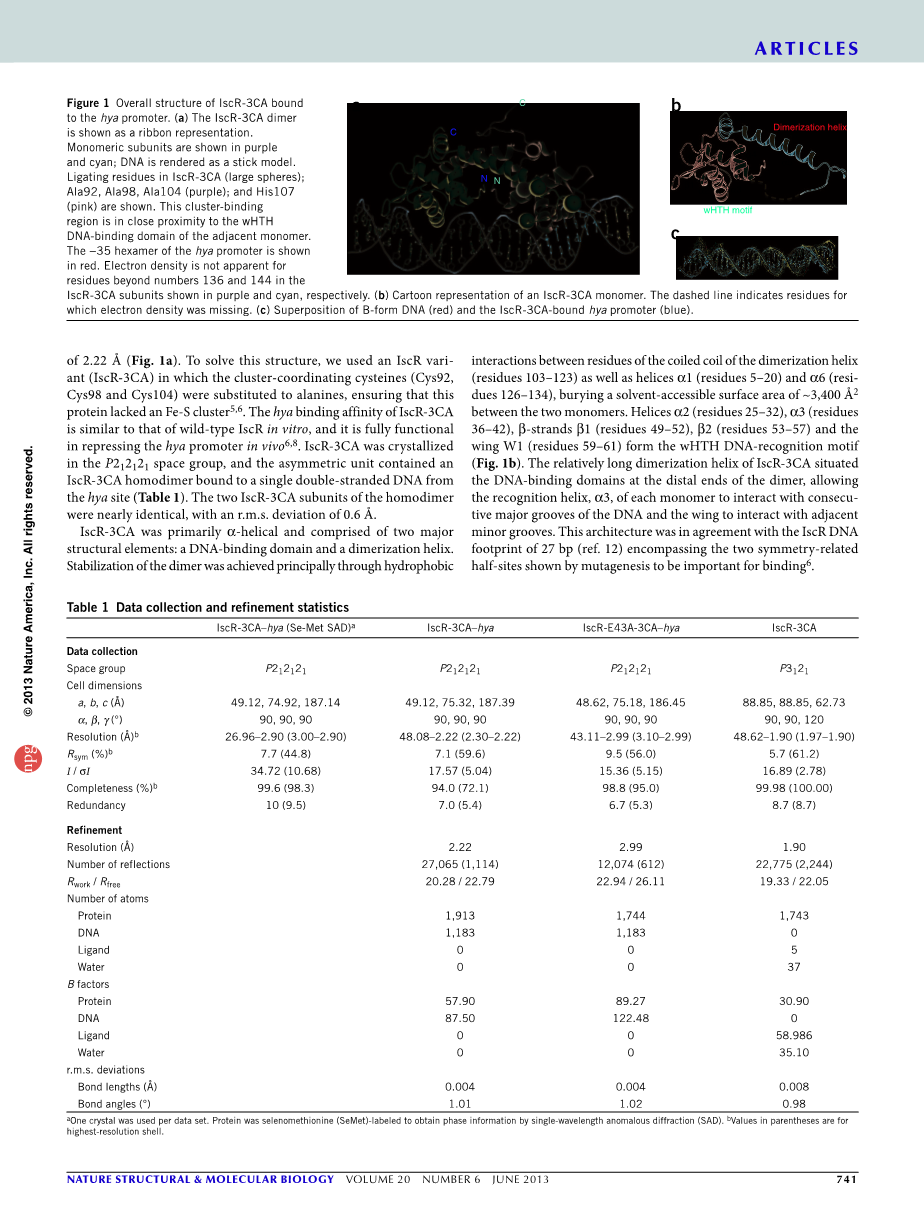
我们利用扩展到分辨率的数据,确定了IscR与含有Hya启动子(IscRhya)2型结合位点的DNA(Hya)的29碱基对(29-bp)片段的晶体结构为2.22A。为了解决这一结构,我们使用了ISCR变体(ISCR-3CA),其中簇协调半胱氨酸(Cys92、Cys98和Cys104)被替换为Alanines,以确保该蛋白缺乏Fe-S簇。在体外,ISCR-3CA的hya结合亲和力与野生型ISCR相似,在体内抑制hya启动子方面具有充分的功能。在P2中结晶ISCR-3CA12121空间群和不对称单元包含一个ISCR-3CA同二聚体,从HYA位点结合到单个双链DNA。同二聚体的两个IscR-3CA亚基几乎相同,具有r.m.s。偏差为0.6A。IscR-3CA主要是alpha;-螺旋结构,由两个主要结构元素组成:DNA结合区和二聚螺旋。二聚体的稳定主要是通过二聚螺旋螺旋(残基103-123)的卷曲线圈残基以及螺旋alpha;1(残基5-20)和alpha;6(残基126-134)之间的疏水相互作用实现的,在两个单体之间掩埋了约34002A的溶剂可及表面积。螺旋alpha;2(残基25–32)、alpha;3(残基36–42)、beta;-链beta;1(残基49–52)、beta;2(残基53–57)和翼W1(残基59–61)形成wHTH DNA识别模体。IscR-3CA相对较长的二聚螺旋位于二聚体末端的DNA结合区,使每个单体的识别螺旋与DNA的连续主槽相互作用,翼与相邻的凹槽相互作用。该体系结构与ISCRDNA足迹27bp一致。包括两个对称相关的半位显示突变是重要的结合。
在野生型IscR中,包含参与[2Fe-2S]簇连接的残基的电子密度在两个蛋白链中只有一个是明显的。电子密度的位置表明这些丙氨酸残基(92、98和104)在这种构象中是溶剂暴露的。如果类似的半胱氨酸残基的位置相似,并且溶剂以配体结合的形式暴露在IscR中,那么这个位置可以允许[2Fe-2S]簇的容易进入和/或丢失,为理解这个蛋白质在感知细胞Fe-Scluster状态中的功能提供了一个框架。另外,来自一个亚单位的簇结合位点靠近另一个亚单位的DNA结合域,表明一个IscR亚单位的Fe-S簇连接可能影响另一个亚单位的构象和DNA结合。
与理想的B型DNA相比,ISCR-3CA结合的HyaDNA弯曲~8.4°。而整体DNA略欠扭曲(HYA为33。1°,而典型B-DNA为35-36°)。将ISCR-3CA的识别螺旋插入主槽,使B型DNA的主槽从11.0A扩大到14.3A核苷酸位置9、10和11。然而,ISCR-3CA-Hya复合物中DNA的总体结构与理想的B-DNA没有明显的差异。
通过apo-IscR识别hya靶点。
我们发现ISCR-3CA具有一个基本的表面,包括识别螺旋和翅膀,它结合DNA。然而,ISCR-3CA-Hya界面的一个显著特征是在这个基本的DNA结合表面的中间存在一个保守的谷氨酸,Glu43,这使得从识别螺旋延伸的侧链羧基与C7C8和C7的外环胺之间发生双齿相互作用a8。酸性残基通常不参与DNA碱基识别,因为具有电负性DNA骨架的静电排斥潜力(尽管有几个例子,包括分解代谢激活蛋白CAPDNA复制引发蛋白RepE-DNA更常见的是锌簇家族中的蛋白质-DNA复合物已报告)。此外,识别螺旋Ser40的羟基与A20和G20的N7形成氢键提出了一个原理,以保护嘌呤在位置20在两个半点的Hya位点。识别螺旋的Gln44侧链。
结构还表明,ISCR-3CA的wHTH结构域的翼深入扩展到hya的小凹槽中,以提供额外的特异性和稳定性。Arg59的侧链延伸到凹槽,其胍基与T6和T25的羰基氧基相互作用。IscR-3CA的hya结合结构提供了对GXXGG模体的深入了解,GXXGG模体在较大的Rrf2家族中,在IscR同源物和蛋白质(如CymR)中都是保守的。Gly60和Gly64被发现存在于其中的构象中只有甘氨酸的Ramachandran空间区域,禁止它们对其他残基的突变,并符合翼内60GXXGG64模体的守恒。此外,该结构表明,Gly60和Gly63必须是甘氨酸的空间原因,因为存在一个C碳会在翼和DNA或DNA之间产生空间冲突分别为1股。值得注意的是,虽然CymR和IscR-3CA之间有相似的非结构化区域。
和在识别螺旋中的三个残基中,有两个在ISCR-3CA和相似的整体褶皱中产生碱基特异性接触,CymR的二聚螺旋的长度和空间取向不同。因此,即使CymR的DNA识别应该与IscR的DNA识别非常相似,CymR结合可能需要蛋白质、DNA或两者的构象变化,正如前面所建议的那样。其他ISCR-3CA-Hya相互作用包括Tyr41,形成a与G18糖基的堆积相互作用和与同一核苷酸的主链磷酸盐的氢键。这种相互作用可能有助于扩大主槽的这个区域的宽度,以稳定识别螺旋的插入。在C7位置,Tyr65与磷酸盐形成氢键和Arg2,Thr4,Ser5,Leu28,Ser38,Arg50和Ser57与磷酸盐骨架进行了额外的接触。
ISCR在hyaDNA结合上的结构变化
为了评估ISCR-3CA与hya启动子结合时的构象变化,我们还在没有DNA的情况下结晶了ISCR-3CA结构在P3中得到2空间组,并使用扩展到1.9A分辨率的数据确定。与以前的报道一致,IscR在溶液中表现为二聚体,ISCR-3CA不结合DNA结晶为同二聚体。游离和hya结合的ISCR-3CA的类似寡聚状态表明,ISCRDNA结合的调控不同于FNR家族的Fe-S转录因子,其中簇诱导的二聚通常控制DNA结合,比较了ISCR-3CA在和DNA结合的形式表明,未结合的ISCR-3CA具有与ISCR-3CA-hya相同的一般结构。二聚体的两个亚基是相对于彼此旋转20°,导致DNA识别螺旋将其定位在主槽中。除了Glu43,它在绑定时旋转,允许它接触C7C8和C7a8只有轻微的旋转位移是必要的,以适应ISCR-3CA与HYA启动子的结合。一般来说,ISCR-3CA翼的残基在DNA结合时几乎没有发生结构变化,只是Arg59的侧链没有明显的电子密度结构,在与Hya的结合后被指挥。参与DNA相互作用的残基结构中IscR-3CA侧链接触的核苷酸碱基与先前被认为对HYA位点突变分析具有重要意义的碱基很好地对应。然而,由于ISCR结合亲和力的降低,突变的hya位点可能是由于核苷酸接触的去除和(或)核苷酸空间阻碍相互作用。
在具有DNA的ISCR侧链中,这些先前的结果可能掩盖了单个核苷酸的相关贡献。为了独立地评估每个侧链-核苷酸的相互作用,我们通过单独地用丙氨酸取代Arg59、Tyr41、Ser40、Gln44或Glu43来去除侧链接触,并了EDTA处理的分离蛋白的结合亲和力,以获得APO形式。我们发现,与野生型蛋白相比,大多数变异的ISCR蛋白与hya位点结合的亲和力降低,表明这些侧链的关键作用。我们观察到Arg59丙氨酸取代之后亲和力降低,表明翼模体的Arg59侧链对结合亲和力作出了主要贡献,部分是通过与富含AT而形成静电相互作用。在那里ge;3bp与狭窄的凹槽的存在相关,这些凹槽通常被精氨酸残基识别。这一结果与以前的调查结果一致这表明,在HYA位点的凹槽AT束内的GC碱基对突变解除了ISCR结合。Tyr41似乎也对DNA结合作出了实质性的贡献,因为在这个位置的丙氨酸取代降低了结合亲和力。这些结果与Tyr41-DNA相互作用一致,提供了结合能,以稳定识别螺旋在主槽。
我们还发现,去除三个ISCR残基的侧链,使碱基特异性相互作用-Ser40,Gln44和Glu43-对DNA结合亲和力有不同的影响。丙氨酸取代Ser40导致结合亲和力适度降低,与Ser40侧链在20和20处识别嘌呤的N7中的作用一致。在HYA位点第20位的嘌呤-拓扑嘧啶取代解除ISCR结合。与T19和T19的Gln44的相互作用似乎没有在与2型模体的结合亲和力中起主要作用,因为Gln44的丙氨酸取代只导致与hya位点结合的缺陷。胸腺嘧啶在19号和19号位置和在2型模体中保守性差,在hya位点的这些位置突变也会导致轻微的结合缺陷。值得注意的是,Glu43的丙氨酸取代对结合亲和力影响不大。尽管利用其羧酸盐侧链与C7C8的N4和C7的N4形成氢键和A8的N6。我们对这一结果感到惊讶,因为hya位点的这些位置的突变显著降低了野生型IscR的结合。
ISCR-E43A失去了对C7C8二核苷酸的鉴别
对IscRE43A变体(IscR)行为的可能解释是消除了Glu43的羧酸盐侧链这些位点的特异性,而不改变结合亲和力。事实上,与野生型ISCR不同,我们发现ISCR-E43A在位置7或8的核苷酸碱基之间没有区分核苷酸碱基,因为半个位点的C7C8突变不像野生型IscR那样破坏结合亲和力,就像它们与野生型ISCR一样。这些结果表明,Glu43接触对识别2型位点的两个半位点中保存良好的CC二核苷酸至关重要。用ISCRE43A和突变型HYA模板进行的附加结合实验表明,尽管对对称的CC二核苷酸失去特异性,但ISCR-E43A在HYA位点的其他保守位置保持与野生型ISCR相同的特异性,并以与野生型ISCR相同的方式与HYA位点结合,如E43Ahya复合物的结构所示。最后,hya的一半位点含有C7′A8′,而不是保守的CC二核苷酸。A8′对胞嘧啶的突变略微提高了结合亲和力(数据未显示),这与Glu43识别的对称CC二核苷酸优先出现在2型位点的两个半位点的位置7和8一致。
ISCR使用Glu43来区分1型模体
与ISCR2型位点相反,1型结合位点是不对称的,在一个半位点和一个T7中含有C7C8二核苷酸t8另一半部位的二核苷酸。因为apo-IscR与2型模体结合,而不是1型模体结合,我们考虑了Glu43与T7的抑制相互作用的可能性t8在1型模体中的二核苷酸阻止APOISCR结合。因此,我们比较了apo-IscR、[2Fe-2S]-IscR、apo-IscR-E43A和[2Fe-2S]-IscR-E43A与来自ISC启动子(ISCRB)的1型位点结合的能力。如预期的那样与[2Fe-2S]-ISCR相比,APO-ISCR结合了ISCRB型1位点。相反,APO-ISCR-E43A强烈地结合了ISCRB型1位点,类似于[2Fe-2S]-ISCR-E43A和野生型[2Fe-2S]-ISCR。这种亲和力
剩余内容已隐藏,支付完成后下载完整资料
资料编号:[416564],资料为PDF文档或Word文档,PDF文档可免费转换为Word
以上是毕业论文外文翻译,课题毕业论文、任务书、文献综述、开题报告、程序设计、图纸设计等资料可联系客服协助查找。
您可能感兴趣的文章
- 纳米材料介导镉污染土壤原位修复的机理及研究进展外文翻译资料
- 纳米零价铁磁洗受镉污染水稻土壤的有效性验证外文翻译资料
- 镉对水稻叶片生长和养分运输的短期影响外文翻译资料
- 间接竞争ELISA的研制及可视化基于金纳米棒生长的多色酶联免疫吸附试验玉米赤霉烯酮的测定外文翻译资料
- 铅(Pb)诱导的小麦根系生化和超微结构变化外文翻译资料
- 用蚕豆根尖微核试验评估污水灌溉区和生物修复区土壤的遗传毒性外文翻译资料
- 镧在红土壤中的生物累积和其对玉米幼苗生长的影响外文翻译资料
- 铈对水稻幼苗生长及部分抗氧化代谢的影响外文翻译资料
- 用黄体孢子菌沉淀碳酸盐固定有毒金属的体外研究及其在含硫化物尾矿中的应用外文翻译资料
- 水牛妊娠早期生物标志物的编码序列克隆及部分序列密码优化后在大肠杆菌中的表达外文翻译资料

